Types of Mutations
The mutation rate for most genes is 1:100,000 to 1:1,000,000. Thus, if an organism has 10,000 genes the chance that any of its gametes would contain a mutant gene is 1 - 10%. There are many recessive mutations that tend to remain hidden in the population until inbreeding (i.e., the mating of individuals more closely related than the average of the population to which they belong) occurs. Inbreeding allows these recessive genes to take on a homozygous condition and become expressed. Some of these genes are lethals, meaning that they will kill the offspring in either the prenatal or postnatal period prior to maturity.
Mutations occur through various events. I will discuss the most common events and those that would be more applicable to cockatiels.
There are several different types of changes that can occur to the DNA structure of a gene that could result in a mutation: a nucleotide (a base, sugar, and phosphate molecule complex) within the DNA strand could be substituted; some of the gene’s DNA could be deleted; or some extra nucleotides could be added to the DNA strand.
Rearrangement of Genetic Material
There are cases where different phenotypes can be obtained when the gene numbers are unequal on the homologous chromosomes.
Moving a gene’s locus could produce a new phenotype. The probability is increased when the gene relocates from euchromatin (lightly stained areas containing genetic information) to near heterochromatin (darkly stained region of the chromosome and assumed to carry little genetic information). The two methods that could move a gene locus are translocation which involves relocating genetic material to a non-homologous chromosome or inversion which involves 180° rotation of a chromosome segment within the same chromosome.
Translocation
When a piece of chromosome breaks off, both ends where the break occurred behave as though they are very receptive to rejoining. When the pieces rejoin into non-homologous combinations, then the event is referred to as translocation. Reciprocal translocation involves the mutual exchange of segments between two non-homologous chromosomes. In the diagram below the effects of a reciprocal translocation are shown.
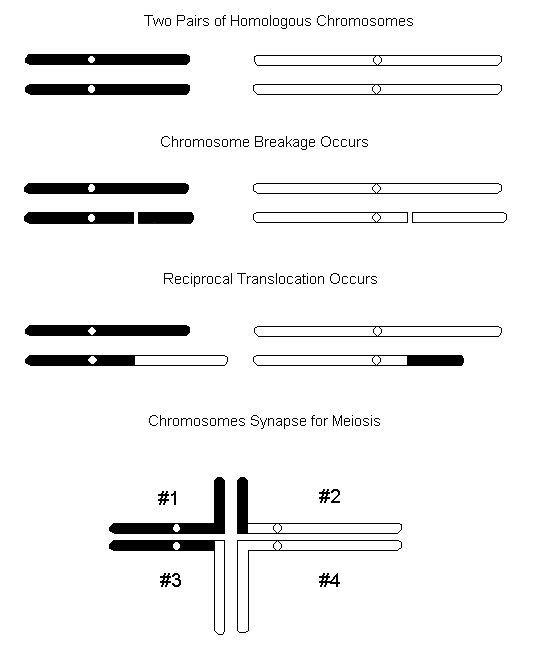
In order to produce genetically balanced and functional gametes, chromosomes #1 and # 4 need to segregate together and chromosomes #2 and #3 need to segregate together. This will not always occur and as a result, non-functional gametes will be produces. In order for functional gametes to be produced, the chromosomes will need to line up in a figure eight pattern during meiotic prophase. This chromosomal pattern has been observed in meiotic prophase tissue of the Northern Green Barbet (Kaul and Ansari 1979).
The translocation heterozygote will display several characteristics: semi-sterility; some genes which used to independently assort will now show linkage; and the phenotypic expression of a gene may be modified since its location has changed.
Translocation has been widely studied in plants. Non-reciprocal translocation has also been observed to occur with human chromosomes. The results of translocation may or may not effect the phenotype. With the above information, I feel it is safe to assume that translocation, either reciprocal or non-reciprocal, occurs to a very small degree with cockatiels. The chromosome with the translocated segment will replicate and pass on its associated phenotype to the offspring.
Inversion
Inversion involves two breaks to a single segment within the same chromosome and the broken segments reinserted into the same chromosome in an inverted position. A plausible explanation for inversion is entanglement of chromosome material during meiotic prophase and simultaneous breakage. See the figure below.
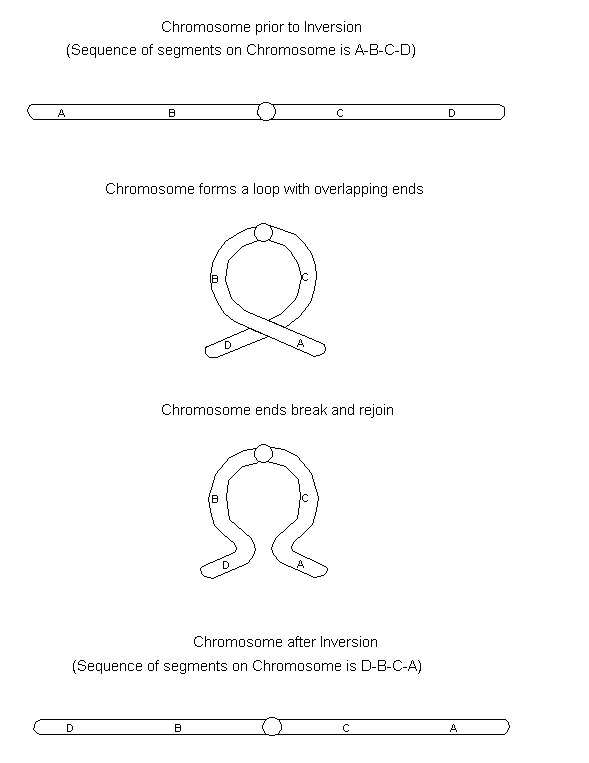
In order for synapsis to occur between the inverted chromosome and its normal homologue, one of the normal homologues will need to loop and the other chromosome will need to wrap around the loop. If crossover was to occur within the inverted segment between these chromosomes in this particular synaptic configuration, gametes containing the crossover product would be non-functional, because they contain deletions or additions.
The inversion heterozygote will, thus, display semisterility and the phenotypic expression of a gene may be modified since its location has changed.
Aneuploidy
Aneuploidy is the term given to variations in the number of chromosomes carried by an individual. Having an extra chromosome (trisomy) can cause a distinctive phenotypic change. A classical example of aneuploidy is Down Syndrome in humans. Down Syndrome occurs when an individual has an extra #21 chromosome. The phenotypic expressions of this syndrome is quite obvious. The gametes produced by a trisomic individual will be 50% normal (only containing one of each chromosome) and 50% abnormal (containing the additional chromosome).
When aneuploidy goes in the opposite direction, that is, loss of a chromosome, the result is typically lethal with the Turner syndrome in humans (XO) being an exception. If the animal does survive, it's fertility is greatly reduced.
Chromosome Deletions
Loss of a large portion of a chromosome may be lethal. However, there are cases where a small portion of a chromosome can be lost and the individual still survives. If a recessive gene exists on the normal chromosome and the dominant allele for this trait exists on the chromosome with the deletion which becomes lost, then the recessive gene will be expressed phenotypically in this hemizygous condition. In humans the cri-du-chat syndrome is associated with deletion of part of the #5 chromosome. This syndrome also has a distinctive phenotypic expression.